Saistītā mantojuma modeļi. Pilnīga un nepilnīga gēnu saite. Hromosomu ģenētisko karšu jēdziens Kas ir pilnīga un nepilnīga gēnu saikne
Saistīto gēnu pārmantošanas mehānismu, kā arī dažu saistīto gēnu atrašanās vietu noteica amerikāņu ģenētiķis un embriologs T. Morgans. Viņš parādīja, ka Mendela formulētais neatkarīgās mantošanas likums ir spēkā tikai gadījumos, kad gēni ar neatkarīgām īpašībām ir lokalizēti dažādās nehomologās hromosomās. Ja gēni atrodas vienā hromosomā, tad pazīmju pārmantošana notiek kopīgi, t.i., saistīti. Šo parādību sāka saukt par saistītu mantojumu, kā arī par saiknes likumu vai Morgana likumu.
Saķeres likums saka: saistītie gēni, kas atrodas vienā hromosomā, tiek mantoti kopā (saistīti). Sajūga grupa- visi gēni vienā hromosomā. Saiknes grupu skaits ir vienāds ar hromosomu skaitu haploīdajā komplektā. Piemēram, cilvēkam ir 46 hromosomas - 23 saišu grupas, zirnī ir 14 hromosomas - 7 saišu grupas, un augļu mušai Drosophila ir 8 hromosomas - 4 saišu grupas. Nepilnīga gēnu saikne- saišu šķērsošanas rezultāts gēni, Tāpēc pilnīga gēnu saikne varbūt organismos, kuru šūnu krustošanās parasti nenotiek.
MORGANA HROMOSOMU TEORIJA. PAMATA NOTEIKUMI.
T. Morgana pētījuma rezultāts bija hromosomu iedzimtības teorijas izveide:
1) gēni atrodas uz hromosomām; dažādas hromosomas satur dažādu skaitu gēnu; katras nehomologās hromosomas gēnu kopums ir unikāls;
2) katram gēnam ir noteikta vieta (lokuss) hromosomā; alēlie gēni atrodas identiskos homologo hromosomu lokusos;
3) gēni atrodas uz hromosomām noteiktā lineārā secībā;
4) gēni, kas lokalizēti vienā hromosomā, tiek mantoti kopā, veidojot sasaistes grupu; saišu grupu skaits ir vienāds ar haploīdu hromosomu kopu un ir nemainīgs katram organisma tipam;
5) šķērsošanas procesā var tikt traucēta gēnu saistība, kas noved pie rekombinanto hromosomu veidošanās; šķērsošanas biežums ir atkarīgs no attāluma starp gēniem: jo lielāks attālums, jo lielāks ir šķērsošanas apjoms;
6) katrai sugai ir unikāls hromosomu kopums – kariotips.
Ar dzimumu saistīts mantojums- Tas ir gēna, kas atrodas uz dzimuma hromosomām, mantojums. Ar Y-hromosomu iedzimtību simptoms vai slimība parādās tikai vīriešiem, jo šīs dzimuma hromosomas sievietes hromosomu komplektā nav. Sievietēm ar X saistīts mantojums var būt dominējošs vai recesīvs, bet vīriešiem tas vienmēr ir, jo ir tikai viena X hromosoma. Ar dzimumu saistīta slimības pārmantošana galvenokārt ir saistīta ar dzimuma X hromosomu. Lielākā daļa iedzimto slimību (noteiktas patoloģiskas īpašības), kas saistītas ar dzimumu, tiek pārnestas recesīvi. Šādu slimību ir ap 100. Sieviete, kura ir patoloģiskas pazīmes nesēja, pati necieš, jo veselīgā X hromosoma dominē un nomāc X hromosomu ar patoloģisko pazīmi, t.i. kompensē šīs hromosomas nepilnvērtīgumu. Šajā gadījumā slimība izpaužas tikai vīriešiem. Ar X saistītais recesīvais tips pārraida: daltonisms (sarkanzaļais aklums), redzes nerva atrofija, nakts aklums, Dišēna tuvredzība, “cirtainu matu” sindroms (rodas traucētas vara vielmaiņas rezultātā, palielināts vara saturs audos, izpaužas kā nedaudz krāsoti, reti un krītoši mati, garīga atpalicība utt.), enzīmu defekts, kas pārvērš purīna bāzes nukleotīdos (kopā ar DNS sintēzes pārkāpumiem Leša-Njēna sindroma veidā, kas izpaužas ar garīgu atpalicību, agresīvs). uzvedība, sevis sakropļošana), hemofilija A (antihemofīlā globulīna - VIII faktora deficīta rezultātā), hemofilija B (Ziemassvētku faktora - IX faktora deficīta rezultātā) utt. Dominējošais X-saistītais tips pārnēsā hipofosfātēmisku rahītu (kuru nevar ārstēt ar vitamīniem D2 un D3), brūnu zobu emalju utt. Šīs slimības attīstās gan vīriešiem, gan sievietēm.
Pilnīga un nepilnīga gēnu saite.
Hromosomu gēniem ir dažādas kohēzijas stiprības. Gēnu saikne var būt: pilnīga, ja rekombinācija nav iespējama starp gēniem, kas pieder vienai un tai pašai saiknes grupai; un nepilnīga, ja ir iespējama rekombinācija starp gēniem, kas pieder tai pašai saiknes grupai.
Hromosomu ģenētiskās kartes.
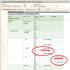
Šīs ir bloķēšanas relatīvās atrašanās vietas diagrammas
iedzimtie faktori – gēni. G.K.H. parādīt reālistiski
esošo lineāro gēnu izvietojuma secību hromosomās (skat. Hromosomu citoloģiskās kartes) un ir svarīgas gan teorētiskajos pētījumos, gan selekcijas darbā, jo ļauj apzināti atlasīt pazīmju pārus krustojumu laikā, kā arī prognozēt dažādu pazīmju pārmantojamības un izpausmes pazīmes pētāmajos organismos. Ja ir G. ch., ir iespējams, mantojot ar pētāmo cieši saistītu “signālgēnu”, kontrolēt to gēnu pārnešanu pēcnācējiem, kas nosaka grūti analizējamu pazīmju attīstību; piemēram, gēns, kas nosaka endospermu kukurūzā un atrodas 9. hromosomā, ir saistīts ar gēnu, kas nosaka samazinātu augu dzīvotspēju.
85. Dzimuma pārmantošanas hromosomu mehānisms. Citoģenētiskās metodes dzimuma noteikšanai.
Stāvs ko raksturo pazīmju komplekss, ko nosaka gēni, kas atrodas hromosomās. Sugām ar divmāju indivīdiem vīriešu un mātīšu hromosomu komplekss nav vienāds, citoloģiski tie atšķiras ar vienu hromosomu pāri, to sauca dzimuma hromosomas. Šī pāra identiskās hromosomas tika sauktas X(x)-hromosomas . Nepāra, nav no otra dzimuma - Y (Y) - hromosoma ; pārējais, kuram nav nekādu atšķirību autosomas(A). Cilvēkiem ir 23 hromosomu pāri. No viņiem 22 autosomu pāri un 1 dzimuma hromosomu pāris. Dzimumu ar identiskām XX hromosomām, kas veido viena veida gametas (ar X hromosomu), sauc homogamētisks, dažāda dzimuma, ar dažādām XY hromosomām, veidojot divu veidu gametas (ar X hromosomu un ar Y hromosomu), - heterogamētisks. Cilvēkiem, zīdītājiem un citiem organismiem heterogamētisks dzimuma vīrietis; putniem un tauriņiem - mātīte.
X hromosomas, papildus gēniem, kas nosaka sieviete, satur gēnus, kas nav saistīti ar dzimumu. Pazīmes, ko nosaka hromosomas, sauc ar dzimumu saistītas īpašības. Cilvēkiem šādas pazīmes ir krāsu aklums (krāsu aklums) un hemofilija (asins nekoagulējamība). Šīs anomālijas ir recesīvas, sievietēm šādas pazīmes neparādās, pat ja šos gēnus pārnēsā kāda no X hromosomām; šāda sieviete ir nēsātāja un nodod tos ar X hromosomu saviem dēliem.
Citoģenētiskā dzimuma noteikšanas metode. Tas ir balstīts uz mikroskopisku hromosomu izpēti cilvēka šūnās. Citoģenētiskās metodes izmantošana ļauj ne tikai pētīt hromosomu normālo morfoloģiju un kariotipu kopumā, noteikt organisma ģenētisko dzimumu, bet, galvenais, diagnosticēt dažādas hromosomu slimības, kas saistītas ar hromosomu skaita izmaiņām. vai to struktūras pārkāpums. Kā ātru metodi, kas nosaka dzimumhromosomu skaita izmaiņas, viņi izmanto Dzimuma hromatīna noteikšanas metode vaiga gļotādas nedalīšanās šūnās. Dzimuma hromatīns jeb Barra ķermenis veidojas sievietes ķermeņa šūnās vienā no divām X hromosomām. Palielinoties X hromosomu skaitam organisma kariotipa sastāvā, tā šūnās veidojas Barra ķermeņi par vienu mazāku daudzumu nekā hromosomu skaits. Kad hromosomu skaits samazinās, ķermeņa nav. Vīriešu kariotipa gadījumā Y hromosomu var noteikt ar intensīvāku luminiscenci, salīdzinot ar citām hromosomām, ja tās apstrādā ar akrihiniprītu un pēta ultravioletajā gaismā.
Hromosomu struktūras iezīmes. Pārmantojamības materiāla organizācijas līmeņi. Hetero- un eihromatīns.
Hromosomu morfoloģija
Mikroskopiskā hromosomu analīze, pirmkārt, atklāj to formas un izmēra atšķirības. Katras hromosomas struktūra ir tīri individuāla. Var arī atzīmēt, ka hromosomām ir kopīgas morfoloģiskās īpašības. Tie sastāv no diviem pavedieniem - hromatīds, atrodas paralēli un savienoti viens ar otru vienā punktā, ko sauc par centromēru vai primāro sašaurinājumu. Dažās hromosomās var redzēt arī sekundāru sašaurināšanos. Tā ir raksturīga iezīme, kas ļauj identificēt atsevišķas hromosomas šūnā. Ja sekundārais sašaurinājums atrodas tuvu hromosomas galam, tad tā ierobežoto distālo reģionu sauc par satelītu. Hromosomas, kas satur satelītu, tiek sauktas par AT hromosomām. Dažās no tām nukleoli veidojas telofāzes laikā.
Hromosomu galiem ir īpaša struktūra, un tos sauc par telomēriem. Telomēriskajiem reģioniem ir noteikta polaritāte, kas neļauj tiem savienoties viens ar otru pārtraukumu laikā vai ar brīviem hromosomu galiem.
Hromatīda (hromosomas) posmu no telomēra līdz centromēram sauc par hromosomas roku. Katrai hromosomai ir divas rokas. Atkarībā no roku garumu attiecības izšķir trīs veidu hromosomas: 1) metacentriskās (vienādas rokas); 2) submetacentrisks (nevienlīdzīgi pleci); 3) akrocentrisks, kurā viens plecs ir ļoti īss un ne vienmēr ir skaidri atšķirams. (p - īsā roka, q - garā roka). Pētījums par hromosomu ķīmisko organizāciju eikariotu šūnās ir parādījis, ka tās galvenokārt sastāv no DNS un olbaltumvielām: histoniem un protomīta (dīgļšūnās), kas veido nukleoproteīnu kompleksu, ko sauc par hromatīnu, kas savu nosaukumu ieguva par spēju krāsoties ar pamata krāsvielas. Olbaltumvielas veido ievērojamu daļu no hromosomu vielas. Tie veido aptuveni 65% no šo konstrukciju masas. Visas hromosomu olbaltumvielas iedala divās grupās: histonos un nehistona proteīnos.
Histoni attēlotas ar piecām frakcijām: HI, H2A, H2B, NZ, H4. Būdami pozitīvi lādēti bāzes proteīni, tie diezgan stingri saistās ar DNS molekulām, kas neļauj nolasīt tajā ietverto bioloģisko informāciju. Tā ir viņu regulējošā loma. Turklāt šie proteīni veic strukturālu funkciju, nodrošinot DNS telpisko organizāciju hromosomās.
Frakciju skaits nehistons olbaltumvielas pārsniedz 100. Starp tiem ir fermenti RNS sintēzei un apstrādei, DNS redublikācijai un labošanai. Hromosomu skābajiem proteīniem ir arī strukturālas un regulējošas funkcijas. Papildus DNS un olbaltumvielām hromosomas satur arī RNS, lipīdus, polisaharīdus un metālu jonus.
Pēc Mendela 1865. gadā noteikto mantošanas noteikumu no jauna atklāšanas 1900. gadā sākās plaša eksperimentāla to piemērojamības pārbaude dažādiem dzīvniekiem un augiem.
Izrādījās, ka ne visi mantojuma gadījumi iekļaujas šajos noteikumos.
Saskaroties vispirms ar atsevišķām novirzēm un pēc tam ar daudziem izņēmumiem no mendelisma, pētnieki tās rūpīgi pētīja. Ģenētikā sāka veidoties cita teorija, plašāka par mendelismu – hromosomu iedzimtības teorija.
Tomass Morgans beidzot saistīja iedzimtības fenomenu ar citoloģiju. Viņš pierādīja, ka iedzimtības materiālie nesēji ir hromosomas, kas atrodas šūnu kodolos ar tajos esošajiem gēniem.
Tā kā katrai sugai ir stingri noteikts hromosomu skaits un dažādu pazīmju skaits ir ļoti liels, jāpieņem, ka vienā hromosomā ir vairāki vai daudzi gēni, kas nosaka pazīmi.
Piemērs: Drosophila ir 1000 gēnu no 4 hromosomu pāriem, 500 gēnu no 10 hromosomu pāriem no kukurūzas, 2000 gēnu no 23 pāriem cilvēkiem, daži zinātnieki apgalvo, ka līdz 1 miljonam gēnu.
Vai katrs gēns var atrasties atsevišķā hromosomā? Nē viņš nevar.
Tāpēc katrā hromosomā ir jābūt daudziem gēniem.
Gametoģenēzes laikā atšķiras hromosomas un līdz ar tām gēni. Gēni, kas atrodas tajā pašā hromosomā, tiek mantoti kā vesela grupa un veido saiknes grupu.
Vienas saišu grupas gēni tiek mantoti neatkarīgi no otras saiknes grupas.
Saiknes grupu skaits ir vienāds ar haploīdu hromosomu kopu.
Piemērs: Drosophila ir 4 hromosomu pāri, no kuriem ir atrastas 4 saiknes grupas; trusis - no 22 hromosomu pāriem atrasti 19 saišu pāri; pele – no 20 hromosomu pāriem atrasti 19 saišu pāri; cilvēkiem - no 23 hromosomu pāriem konstatētas 25 saišu grupas, 22 grupas - pēc autosomu pāru skaita, X un Y hromosomās un 25. saišu grupa mitohondriju DNS.
Saistītās īpašību pārmantošanas fenomenu 1906. gadā atklāja Bateson un Punett. Šie pētnieki pētīja saldo zirņu augu krustošanu, kas atšķīrās pēc divām īpašībām: ziedputekšņu formas un ziedu krāsas. Saskaņā ar Mendeļa likumiem otrās paaudzes hibrīdos šajā gadījumā ir jāievēro dihibrīda krustojumam raksturīgo īpašību sadalījums 9: 3: 3: 1.
Tomēr Batsons un Punnets konstatēja citādi. Šķiet, ka hibrīdos divas rakstzīmes (ziedputekšņu forma un ziedu krāsa) centās palikt sākotnējās vecāku kombinācijās.
Batsons un Punnets nespēja to izskaidrot.
Izskaidrojumu šai parādībai vēlāk sniedza Tomasa Morgana skola, kas ieviesa terminu gēnu saite. Viņš pierādīja, ka gēni, kas atrodas vienā hromosomā, ir cieši saistīti viens ar otru, t.i. savienoti un sakārtoti lineārā secībā.
Radījis hromosomu iedzimtības teoriju, Morgans pierādīja, ka pastāv pilnīga un nepilnīga gēnu saikne.
Morgan veica savus eksperimentus ar augļu mušu.
Pirmajam eksperimentam tika ņemtas mušas: tēviņi ar pelēku ķermeni un rudimentāriem spārniem, kurus viņš krustoja ar mātītēm ar melnu ķermeni un gariem spārniem.
d koncepcija D garums
Gametes C c
C d – gēni atrodas vienā hromosomā. Tie ir homozigoti abiem pāriem. Hibrīdiem F 1 bija 100% pelēks ar gariem spārniem (heterozigots). Viņš atlasīja tēviņus no F 1 un krustoja tos ar mātītēm, kurām bija 2 recesīvās pazīmes (testa krustojums).
♂ С с ♀ с
d D d ieņemts
pelēks melns
garš rudimentārs
Ar ar ar ar In F parādījās 2. paaudze
2 veidu pēcnācēji:
50% - pelēks ar rudimentāriem spārniem,
d d D d 50% - melns ar gariem spārniem.
pelēks melns
ieņemts garums
Ir notikusi pilnīga gēnu saistība.
II pieredze. Morgans paņēma mātītes no F 1 pelēkajiem garajiem spārniem.
♀ ssdd x ♂ ssdd (analizē)
pelēks melns
garums ieņemts
S s s s s s
d d D d d d d
pelēks melns melns pelēks
ieņemts garums ieņemts garums
Otrajā paaudzē iegūti 4 dažādi fenotipi: 145 – melni garspārni = 41,5%; 150 – pelēki rudimenti = 41,5%.
Tāpat kā vecāku veidlapas:
28% - pelēki garie spārni - 8,5%
33% - melns rudimentārs - 8,5%
jaunas kombinācijas 17%
Šajā gadījumā sajūgs nebija pilnīgs. Tomēr gēni, kas atrodas tajā pašā hromosomā, nav absolūti saistīti. Lielākajai daļai indivīdu ir vecāku īpašības, un nelielai daļai indivīdu ir jaunas rakstzīmju kombinācijas. Nepilnīgas sakabes iemesls ir šķērsošana.
Kā var izskaidrot šo jaunu kombināciju fenomenu, ja gēni ir vienas hromosomas daļa?
Tas izskaidrojams ar to, ka gametoģenēzes laikā (reducēšanas dalīšanās laikā) hromosomas apvij viena ap otru, apmainās ar daļām, un pēc tam tās atdalās (lūzt), radot jaunas hromosomas (viena daļa no mātes, otra no tēva).
Šķērsojot
Hromosomu sekciju apmaiņas procesu sauc - hromosomu šķērsošana vai šķērsošana.
Šķērsošanas mehānisma klātbūtne paplašina kombinētās mainīguma iespēju, un tai ir liela nozīme dzīvnieku pasaules evolūcijā.
Krustošana notiek, kad gēni ir heterozigotā stāvoklī. Indivīdi, kas iegūti ar krustojuma palīdzību, tiek saukti par krosoveriem un bez šķērsošanas - nekrosoveriem.
Šķērsošana var būt viena, divvietīga vai trīskārša. Visbiežāk vienvietīgi, retāk divvietīgi un trīsvietīgi. Tas izskaidrojams ar to, ka hromosoma ir elastīgs ķermenis, kura dēļ cilpas veidošanās vienā apgabalā kavē tās veidošanos citā, kā rezultātā retāk notiek šķērsošana divās zonās. Bremzēšanas fenomenu sauc iejaukšanās. Ar dubultu šķērsošanu hromosomas tiek sadalītas divos punktos, kā rezultātā tās apmainās ar viduspunktiem; ar trīskāršu šķērsošanu - trīs punktos ar divu hromosomu sekciju apmaiņu, kas izraisa vēl lielāku hromosomu mainīguma pieaugumu. dzimumšūnas. Tomēr dubultie un trīskāršie krosoveri notiek daudz retāk nekā atsevišķi krosoveri.
Lielākajā daļā augu un dzīvnieku sugu hromosomu skaits (2n) nepārsniedz vairākus desmitus, un gēnu skaits var sasniegt desmitus un pat simtus tūkstošus. Tas nozīmē, ka vienā hromosomā var koncentrēties tūkstošiem gēnu, un tiem ir jābūt mantotiem kopā, tas ir, saistītiem. Šo mantojumu sauc "gēnu saikne". Gēni, kas atrodas hromosomā, veido saiknes grupu. Sugai ir tik daudz šādu saišu grupu, cik tai ir hromosomu pāru. Ja gēni A un B atrodas dažādos hromosomu pāros, tad, krustojot genotipus ♀AABBx♂aabb otrajā paaudzē, ievērojot abu gēnu pilnīgu dominēšanu, tipisks atkausēšanas modelis pēc fenotipa (9: 3: 3: 1) parādīsies. Gametoģenēzes laikā AaBb genotipa indivīds ražo 4 veidu gametas: AB, Ab, aB, ab.
Ja gēni A un B atrodas ūdens hromosomā, tad, šķērsojot genotipus ♀ABABx♂abab, F2 rodas šķelšanās, kas līdzīga sadalīšanai monohibrīda krustojumā: 1ABAB:2ABab:1abab. Tas notiek tāpēc, ka F1 hibrīdi veido 2 veidu gametas: AB, ab. Šajā gadījumā tiek novērota pilnīgas gēnu saiknes parādība. Izmantojot augļu mušu Drosophila melanogaster, ir atrisinātas daudzas sarežģītas ģenētikas problēmas - mākslīga mutāciju veidošanās, gēna būtība, genoma atkodēšana utt.
Drosophila ir maza izmēra (apmēram 3 mm gara), pelēkā krāsā, ar spilgti sarkanām acīm (3.1. att.). Paši dienvidi ir lielāki nekā tēviņi. Viņu viss vēders ir svītrains, un beigās tas ir smails (atrodas olšūna). Tēviņiem pēdējie segmenti ir sapludināti, un zirga vēders ir pilnīgi tumšā krāsā un noapaļots.

Savvaļā sastopams daudzos kontinentos. Vasarā ar augļu un dārzeņu plūsmu no mūsu valsts dienvidu reģioniem tas iekļūst tālu uz ziemeļiem. Drosophila barojas ar augļiem, dārzeņiem un koku sulām.
Laboratorijas apstākļos Drosophila audzē stikla traukos uz īpašas barotnes 25 °C temperatūrā. Tās attīstības cikls no apaugļotas olšūnas līdz pieaugušajam ir aptuveni 10 dienas. Gada laikā jūs varat iegūt apmēram 40 Drosophila paaudzes.
Drosophila diploīdais komplekts sastāv no 8 hromosomām. Otrais hromosomu pāris satur melno (melnā ķermeņa krāsa - b) un vestigiālo (rudimentārie spārni - vg) gēnus. Dominējošās savvaļas tipa alēles b+ izraisa pelēku ķermeņa krāsojumu un normālus vg-spārnus. Pelēkās ķermeņa krāsas b+ un rudimentāro spārnu vg (recesīvā alēle) gēni atrodas vienā hromosomā, melnās krāsas b (recesīvā alēle) un garo spārnu gēni vg+ atrodas citā homologā hromosomā. Krustojot pelēkus rudimentārus spārnu tēviņus (b+vg) ar melnajām garspārnajām mātītēm (bvg-), F1 hibrīdi saņem no tēva hromosomu ar pelēkās krāsas un rudimentāro spārnu gēniem (3.2. att.).
Pēc tam tika atklāts gēnu nepilnīgas sasaistes fakts (3.3. att.). Amerikāņu ģenētiķis T. Morgans Drosofilā pētīja nepilnīgu gēnu saikni un šķērsošanu. Drosophila tēviņiem krustošanās nenotiek. Ņemsim mātītes ar b+vg+bvg genotipu un tēviņus ar bvgbvg genotipu. Ja mātītēm krustošanās nenotiek, tad iegūst divas fenotipiskās klases. Ja notiek krustošanās un alēles maina vietas hromosomu pārī, tiek iegūtas divas papildu fenotipiskās klases. 1% krosoveru fenotipu tiek ņemti par šķērsošanas vienību. Viņa tika nosaukta par Morganidu. Pašlaik tas ir apzīmēts kā centimorgans (sM).
3.3. attēlā redzams, ka attālums starp gēniem b un vg ir 17 centimorgans.
Pētot gēnu krustošanos, tika noteikta gēnu relatīvā atrašanās vieta. Ja krustošanās biežums starp gēniem A un B ir 12%, starp B un C ir 8%, bet starp A un C ir 20%, tad gēns B atrodas starp gēniem A un C. Turklāt tika atklāts, ka gēni atrodas hromosomā lineārā secībā. Pamatojoties uz to, kļuva iespējams apkopot hromosomu ģenētiskās kartes un noteikt gēnu atrašanās vietu.
Izmantojot hromosomu ģenētiskās kartes, ir iespējams paredzēt šo organismu pētīto īpašību mantojuma raksturu un, izmantojot selekcijas darbā, veikt apzinātu pāru atlasi krustošanai.
Galvenie hromosomu iedzimtības teorijas nosacījumi ir šādi:
1) gēni atrodas uz hromosomām un ir izkārtoti lineāri;
2) gēni, kas atrodas ūdens hromosomā, pieder vienai un tai pašai saišu grupai. Saiknes grupu skaits ir vienāds ar haploīdu hromosomu skaitu;
3) pazīmes, kuru gēni atrodas vienā hromosomā, ir iedzimtas saistītas;
4) pamatojoties uz gēnu lineāro izvietojumu hromosomās un krustošanās biežumu, kā attāluma starp gēniem indikatoru, var izveidot hromosomu kartes.
Gēni, kas atrodas tajā pašā hromosomā, ir sajūga grupa. Gēnu saikne- Tas ir vienā un tajā pašā hromosomā esošo gēnu kopīgs mantojums. Saiknes grupu skaits atbilst haploīdajam hromosomu skaitam. Gēnu saikne, kas atrodas tajā pašā hromosomā, var būt pilnīga vai nepilnīga. Pilna saķere: Morgans krustoja melnas garspārnu mātītes ar pelēkiem tēviņiem ar rudimentāriem spārniem. Drosofilā pelēkā ķermeņa krāsa dominē pār melnu, un garie spārni dominē pār rudimentārajiem spārniem. Pelēks korpuss - A, melns korpuss a; garspārni - B, rudimentāri spārni - c. Spermioģenēzes laikā meiozes laikā homologās hromosomas sadalās dažādās dzimumšūnās. 1) AA//AB*av//av=4AB//av; 2) AB//av*AB//av=AB//AB, AB//av, ab//AV, ab//av. Ja gēni ir atrodami autosomās, tad ar pilnīgu saikni F1 būs fenotipa vienveidība, bet F2 - 3:1 neatkarīgi no tā, cik daudzās pazīmēs vecāki atšķiras, jo tiek pētīts viens hromosomu pāris.
Nepilnīgas saiknes parādība pazīmju pārmantošanā
Krustošanas rezultātā pēcnācējiem bija īpašību kombinācija, tāpat kā sākotnējās vecāku formas, bet indivīdi parādījās arī ar jaunu pazīmju kombināciju - sajūgs nepilnīgs. B – pelēks, c – melns, V – normāls, v – rudimentārs. Bv||Bv*bV||bV=Bv||bV; mātītes no pirmās paaudzes tika krustotas ar analizatora tēviņiem: BV//bV*bv//bv=Bv//bv,bV//bv – nav crossover. Bv//bV*bv//bv=2bv//bv, 2BV//bv – krosovers. Homoloģisko hromosomu apmaiņu ar to daļām sauc krustojums vai šķērsošana. Tiek izsauktas personas ar jaunām rakstzīmju kombinācijām, kas izriet no krustošanās krosoveri. Jaunu formu parādīšanās skaits ir atkarīgs no krustošanās biežuma, ko nosaka pēc šādas formulas: Krosojuma biežums = (Krustošanas formu skaits) 100/ Kopējais pēcnācēju skaits. Tiek pieņemts, ka krusta mērvienība ir vienāda ar 1%. Viņu sauc par morganidu. Krosoveru apjoms ir atkarīgs no attāluma starp pētāmajiem gēniem. Jo tālāk gēni atrodas viens no otra, jo biežāk notiek krustojums; jo tuvāk tie atrodas, jo mazāka iespēja šķērsot.
Hromosomu kartes. To uzbūves piemērs
Hromosomu karte- gēnu atrašanās vietas plāns hromosomā. Gēni atrodas hromosomās lineārā secībā noteiktos attālumos viens no otra. Tiek saukts fenomens, kas kavē šķērsošanu vienā apgabalā, šķērsojot citā iejaukšanās. Jo mazāks attālums, kas atdala trīs gēnus, jo lielāka ir iejaukšanās. Ņemot vērā gēnu lineāro izvietojumu hromosomā, kā attāluma vienību ņemot frekvenču šķērsošanu, Morgans sastādīja pirmo gēnu atrašanās vietas karti vienā no Drosophila hromosomām: сh___13.6___ y___28.2___b. Veidojot kartes, tās nenorāda attālumu starp gēniem, bet gan attālumu līdz katram gēnam no hromosomas sākuma nulles punkta. Dominējošā alēle ir apzīmēta ar lielo burtu, bet recesīvā alēle - ar mazo burtu. Pēc ģenētisko karšu izveidošanas radās jautājums, vai gēnu atrašanās vieta hromosomā, kas konstruēta, pamatojoties uz krustošanās frekvenci, atbilst patiesajai atrašanās vietai. Katrai hromosomai visā tās garumā ir specifiski disku modeļi, kas ļauj atšķirt tās dažādās sadaļas vienu no otras. Pārbaudes materiāls bija hromosomas, kurās mutācijas rezultātā radās dažādas hromosomu pārkārtošanās: trūka atsevišķu disku vai tie bija apgriezti, vai dubultojās. Fiziskie attālumi starp gēniem ģenētiskajā kartē ne visai atbilst noteiktajiem citoloģiskajiem attālumiem. Tomēr tas nemazina hromosomu ģenētisko karšu vērtību, lai prognozētu indivīdu iespējamību ar jaunām pazīmju kombinācijām. Pamatojoties uz daudzu Drosophila eksperimentu rezultātu analīzi, T. Morgans formulēja hromosomu teorija iedzimtība, kuras būtība ir sekojoša: 1) gēni atrodas hromosomās, kas atrodas tajās lineāri noteiktā attālumā viens no otra; 2) gēni, kas atrodas vienā hromosomā, pieder vienai un tai pašai saišu grupai. Saiknes grupu skaits atbilst haploīdajam hromosomu skaitam; H) pazīmes, kuru gēni atrodas vienā hromosomā, ir iedzimtas saistītas; 4) heterozigotu vecāku pēcnācējiem mejozes procesa laikā var rasties jaunas gēnu kombinācijas, kas atrodas vienā hromosomu pārī. Šķērsošanas biežums ir atkarīgs no attāluma starp gēniem; 5) pamatojoties uz gēnu lineāro izvietojumu hromosomā un šķērsošanas biežumu kā attāluma starp gēniem indikatoru, var izveidot hromosomu kartes.
Biseksualitāte, interseksualitāte, giandromorfisms, himērisms pēc dzimuma hromiem. Hormonu un vides apstākļu nozīme dzimumpazīmju attīstībā
Jebkurai zigotai ir x-hromi un autosomas, t.i. ir gan sieviešu, gan vīriešu gēni, t.i. ģenētiski jebkurš organisms biseksuāls(biseksuāls). Intersekss– hermafrodīti – indivīdi ar attīstītām gan sieviešu, gan vīriešu īpašībām. 2 veidi: patiess - ir sieviešu un vīriešu dzimumdziedzeri gēnu nelīdzsvarotības dēļ; nosacīti - ir viena dzimuma dziedzeri un otra dzimuma ārējās seksuālās īpašības hormonu nelīdzsvarotības dēļ. Dažreiz atrodams kukaiņos un dzīvniekos giandromorfi– vienai ķermeņa daļai ir sievietes, bet otrai vīrišķās īpašības. Cēloņi: Sieviešu zigota ir sadalīta 2 blastomēros. Viens no viņiem zaudēja vienu x-lame. No šī blastomēra veidosies vīrišķā ķermeņa puse. Himērisms puse no hromosomām xx/xy ir atrodamas daudzdzimušiem dzīvniekiem, buļļiem - kad viens un tas pats organisms satur xx hromosomas, un ir traucēta xy hromosomu vairošanās. Pie normālas barošanas tēviņi aug, un, ja pārtikai pievieno sievišķos dzimumhormonus, aug mātītes (zivju mazuļi). Ja jūras tārpa kāpurs pievienojas jūras dibenam, tā ir mātīte, ja tā pieķeras mātītes cilpiņai, tas ir tēviņš.
Dzimuma noteikšanas veidi dzīvniekiem. Primārā un sekundārā dzimuma attiecība. Dzimumu regulējuma problēma
Apņēmība nodrošina vienāda skaita tēviņu un mātīšu veidošanos, kas nepieciešama sugas normālai pašvairošanai. Veidi: 1) epigāms - indivīda dzimums tiek noteikts ontoģenēzes laikā un ir atkarīgs no ārējās vides. 2) progamous - dzimums tiek noteikts gametoģenēzes laikā indivīda vecākiem. 3) singāms - dzimums tiek noteikts gametu saplūšanas brīdī. Primārā un sekundārā dzimuma attiecība: dzimumu attiecība, kaķis tiek noteikts gametu saplūšanas brīdī, sauc primārs, vienmēr 1:1. Tiek sauktas jebkuras dzimumu attiecības izmaiņas pirms vai pēc dzimšanas sekundārais. Parasti pēc piedzimšanas tas pāriet par labu sieviešu dzimumam, tāpēc daudzām dzīvnieku sugām un cilvēkiem piedzimst vairāk tēviņu nekā mātīšu: truši - 57%, cilvēki - 51%, putni - 59%. Dzimumu regulēšanas problēma: ir liela ekonomiska nozīme. Piemēram: piena lopkopībā un dējējputnu audzēšanā ir vēlamas mātītes, un tur, kur galvenais produkts ir gaļa, labāk ir tēviņi. Problēma ir sadalīt spermu x un y frakcijās. Metodes: 1) elektroforēze - x - spermatozoīdiem ir negatīvs lādiņš - virzās uz katodu, un y - spermatozoīdi virzās uz anodu. 80% garantija. 2) Sedimentācijas metode - x - sperma ir blīvāka un nosēžas, un y - paliek virspusē. 3) Izmantojot skābju komplektu, lai mainītu sievietes reproduktīvā trakta pH, lai radītu apstākļus tikai x - vai tikai y-. 4) Partenoģenēze: genoģenēze - mātīšu iegūšana - olšūnu apstaro ar rentgena stariem. pirmajā kārtā, tādējādi aizkavējot hroma-m novirzi, veidojas olšūna ar diploīdu hroma-m komplektu, un mātīte bez apaugļošanas kļūst par kaķi. Androgēze - tēviņu iegūšana - olšūnas kodols tiek nogalināts ar rentgena stariem, tad tajā iekļūst divi spermatozoīdi, kodoli saplūst, dodot diploīdu komplektu, būs tēviņš. 5) Spermas sadalīšanas paņēmiens frakcijās, pamatojoties uz DNS daudzumu spermā. 6) Jo jaunāki vecāki, jo lielāka iespēja, ka viņi piedzims kā vīrieši. 7) Jo vairāk spermatozoīdu sieviešu dzimumorgānos, jo lielāka iespējamība, ka piedzims vīrietis. 8) Jo ilgāk tiek uzglabāta sperma – mātīte. 9) Mājputnu barošana: ja pievienojat Ca gaiļa barībai, tas ir mātīte, un, ja K, tas ir tēviņš. 10) Jebkurā populācijā ir spēkā līdzsvara likums, t.i. Dzimumu attiecība mēdz būt 1:1.
1906. gadā V. Batsons un R. Punnets, krustojot saldo zirņu augus un analizējot ziedputekšņu formas un ziedu krāsas pārmantošanu, atklāja, ka šīs īpašības nedod neatkarīgu izplatību pēcnācējos, hibrīdi vienmēr atkārto vecāku formu īpašības. Kļuva skaidrs, ka ne visas pazīmes raksturo neatkarīga izplatība pēcnācējos un brīva kombinācija.
Katram organismam ir milzīgs skaits īpašību, bet hromosomu skaits ir mazs. Līdz ar to katrā hromosomā ir nevis viens gēns, bet vesela gēnu grupa, kas ir atbildīga par dažādu pazīmju attīstību. Viņš pētīja to pazīmju pārmantošanu, kuru gēni ir lokalizēti vienā hromosomā. T. Morgans. Ja Mendels veica savus eksperimentus ar zirņiem, tad Morganam galvenais objekts bija augļu muša Drosophila.
Drosophila rada daudzus pēcnācējus ik pēc divām nedēļām 25 °C temperatūrā. Pēc izskata vīrietis un mātīte ir skaidri atšķirami – tēviņam ir mazāks un tumšāks vēders. Viņiem ir tikai 8 hromosomas diploīdā komplektā, un tie diezgan viegli vairojas mēģenēs uz lētas barotnes.
Krustojot Drosophila mušu ar pelēku ķermeni un normāliem spārniem ar mušu ar tumšu ķermeņa krāsu un rudimentāriem spārniem, pirmajā paaudzē Morgans ieguva hibrīdus ar pelēku ķermeni un normāliem spārniem (dominē gēns, kas nosaka vēdera pelēko krāsu). tumšā krāsa un gēns, kas nosaka normālu spārnu attīstību, - virs mazattīstītu spārnu gēna). Veicot F 1 mātītes analītisko krustošanu ar tēviņu, kuram bija recesīvās pazīmes, teorētiski bija paredzēts iegūt pēcnācējus ar šo pazīmju kombinācijām attiecībā 1:1:1:1. Tomēr pēcnācējos nepārprotami dominēja indivīdi ar vecāku formu pazīmēm (41,5% - pelēki garspārni un 41,5% - melni ar rudimentāriem spārniem), un tikai nelielai daļai mušu bija dažādu raksturu kombinācijas, kas atšķiras no vecāki (8,5% - melni garspārni un 8,5% - pelēki ar rudimentāriem spārniem). Šādus rezultātus varētu iegūt tikai tad, ja gēni, kas ir atbildīgi par ķermeņa krāsu un spārnu formu, atrodas vienā hromosomā.

1 - nešķērsojošās gametas; 2 - krusteniskās gametas.
Ja ķermeņa krāsas un spārnu formas gēni ir lokalizēti vienā hromosomā, tad šīs krustošanās rezultātā divām indivīdu grupām bija jāatkārto vecāku formu īpašības, jo mātes organismam vajadzētu veidot tikai divu veidu gametas - AB un ab, un tēva viens - viens tips - ab . Līdz ar to pēcnācējos jāveido divas indivīdu grupas ar genotipu AABB un aabb. Tomēr pēcnācējos parādās indivīdi (kaut arī nelielā skaitā) ar rekombinētām rakstzīmēm, tas ir, ar genotipiem Aabb un aaBb. Lai to izskaidrotu, jāatgādina dzimumšūnu veidošanās mehānisms – mejoze. Pirmā meiotiskā dalījuma fāzē homologās hromosomas tiek konjugētas, un šajā brīdī starp tām var notikt reģionu apmaiņa. Šķērsošanas rezultātā atsevišķās šūnās notiek hromosomu sekcijas apmaiņa starp gēniem A un B, parādās gametas Ab un aB, kā rezultātā pēcnācējos veidojas četras fenotipu grupas, tāpat kā ar brīvu kombināciju. gēni. Bet, tā kā krustošanās notiek nelielas gametu daļas veidošanās laikā, fenotipu skaitliskā attiecība neatbilst attiecībai 1:1:1:1.
Sajūga grupa- gēni, kas lokalizēti vienā hromosomā un mantoti kopā. Saiknes grupu skaits atbilst haploīdajam hromosomu kopumam.
Ķēdes mantojums- tādu pazīmju pārmantošana, kuru gēni ir lokalizēti vienā hromosomā. Saiknes stiprums starp gēniem ir atkarīgs no attāluma starp tiem: jo tālāk gēni atrodas viens no otra, jo lielāks ir krustošanās biežums un otrādi. Pilna saķere- saistīts mantojuma veids, kurā analizējamo pazīmju gēni atrodas tik tuvu viens otram, ka to krustošanās kļūst neiespējama. Nepilnīgs sajūgs- saistīts mantojuma veids, kurā analizējamo pazīmju gēni atrodas noteiktā attālumā viens no otra, kas padara iespējamu krustošanos starp tām.
Neatkarīgs mantojums- tādu pazīmju pārmantošana, kuru gēni ir lokalizēti dažādos homologo hromosomu pāros.
Gametas, kas nav krusteniskas- gametas, kuru veidošanās laikā šķērsošana nenotika.
Nerekombinanti- hibrīdi indivīdi, kuriem ir tāda pati īpašību kombinācija kā viņu vecākiem.
Rekombinanti- hibrīdi indivīdi, kuriem ir atšķirīga īpašību kombinācija nekā viņu vecākiem.
Attālums starp gēniem tiek mērīts Morganids— konvencionālās vienības, kas atbilst krustenisko gametu procentuālajai daļai vai rekombinanto šūnu procentuālajai daļai. Piemēram, attālums starp pelēkās ķermeņa krāsas gēniem un gariem spārniem (arī melnu ķermeņa krāsu un rudimentāriem spārniem) Drosofilā ir 17% jeb 17 morganīdu.
Diheterozigotos dominējošie gēni var atrasties vai nu vienā hromosomā ( cis fāze), vai citā ( transfāze).

1 - Cis-fāzes mehānisms (ne-crossover gametas); 2 - transfāzu mehānisms (ne-crossover gametas).
T. Morgana pētījumu rezultāts bija radīšana hromosomu iedzimtības teorija:
- gēni atrodas hromosomās; dažādas hromosomas satur dažādu skaitu gēnu; katras nehomologās hromosomas gēnu kopums ir unikāls;
- katram gēnam ir noteikta vieta (lokuss) hromosomā; alēlie gēni atrodas identiskos homologo hromosomu lokusos;
- gēni atrodas uz hromosomām noteiktā lineārā secībā;
- gēni, kas lokalizēti vienā hromosomā, tiek mantoti kopā, veidojot saiknes grupu; saišu grupu skaits ir vienāds ar haploīdu hromosomu kopu un ir nemainīgs katram organisma tipam;
- šķērsošanas laikā var tikt traucēta gēnu saite, kas noved pie rekombinanto hromosomu veidošanās; šķērsošanas biežums ir atkarīgs no attāluma starp gēniem: jo lielāks attālums, jo lielāks ir šķērsošanas apjoms;
- Katrai sugai ir unikāls hromosomu komplekts - kariotips.
Iet uz lekcijas Nr.17“Ģenētikas pamatjēdzieni. Mendeļa likumi"